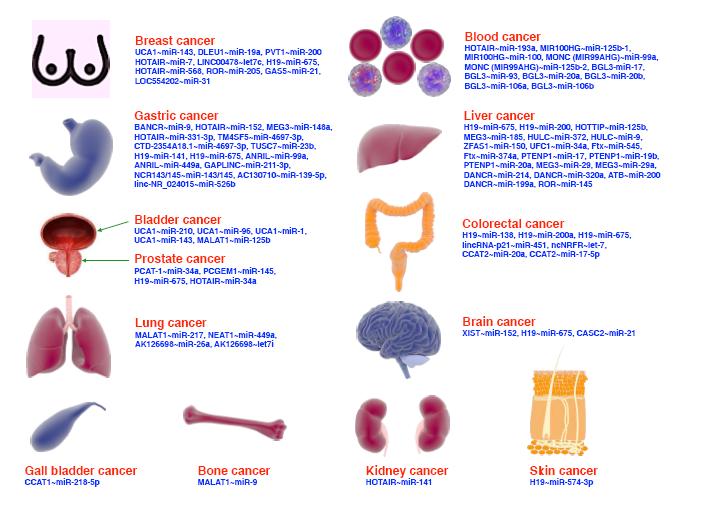

活性氧族是一类氧衍生的代谢物,被广泛地认为是多种生理过程以及病理状态下关键的调节剂,在血管系统中主要由还原型辅酶Ⅱ氧化酶生成。慢性创口的愈合涉及2种不同形式的血管新生:血管发生(骨髓来源分化而成的循环内皮祖细胞形成)和血管生成(已存在血管局部内皮细胞的芽生而形成)。活性氧族通过对血管新生过程中所涉及的内皮祖细胞、内皮细胞和血管平滑肌细胞功能的调节,影响创口愈合。
引用本文: 谢飞, 何延政, 刘勇. 活性氧簇对创口愈合过程中血管新生的影响. 华西医学, 2014, 29(10): 1979-1982. doi: 10.7507/1002-0179.20140599 复制
活性氧簇(ROS)是一类氧衍生的具有较强化学反应的活性分子,包括超氧阴离子、过氧化氢、过氧亚硝基阴离子、次氯酸、羟自由基、一氧化氮等。它的生成是多种刺激因素作用的结果,这些刺激因素包括生长因子、细胞因子、低氧诱导因子、剪切应力等。传统观点认为,ROS通过自由基链式反应引起生物体的核酸、脂肪、蛋白质等大分子的损害。近几十年来,大量的研究表明,氧化应激对于血管疾病的发病具有重要的作用,例如与高血压、动脉粥样硬化和再狭窄的发生有着密切的关系。近年的研究也表明,对于多种生理活动过程而言,ROS均具有重要的调节作用,细胞内多种重要信号路径的调节都是由ROS介导的,例如G蛋白偶联受体、Notch路径[1]、Wnt-β-catenin路径[2]、促分裂原活化蛋白激酶路径、JAK-STAT路径、核因子κB(NF-κB)、PI3K/AKT路径等,其调节信号的转导过程被称之为可逆性的氧化还原过程。除此之外,ROS还可以激活转录因子(例如激活蛋白1、NF-κB自身、c-Jun氨基末端激酶等),调控细胞的生长、分化、迁移、凋亡、衰老等多种生理活动。
创口愈合缓慢会增加医疗负担,尤其影响老年、糖尿病及肥胖患者。组织损伤及缺血所引起的血管生长对于创口的愈合非常关键。目前发现存在2种不同形式的血管新生:血管的发生,由骨髓来源分化而成的循环内皮祖细胞(EPC)所形成;血管的生成,由已存在血管局部内皮细胞(EC)的芽生而形成。创口血管的新生包括血管的发生、生成以及血管平滑肌细胞(VSMC)迁移从而形成血管壁的过程[3]。这一过程涉及EPC的动员和分化、EC的迁移、VSMC表型的转化等,ROS在其中的作用不可或缺。因此,现就ROS如何影响创口愈合过程中血管的新生进行综述。
1 ROS的来源与还原型辅酶Ⅱ氧化酶家族(Nox)
ROS来源于包括Nox、黄原呤氧化酶、线粒体呼吸链、非耦合的一氧化氮合成酶、髓过氧化物酶、脂氧合酶、环氧合酶、细胞色素P450和血红素加氧酶的多酶系统。在上述多种能够产生ROS的氧化酶中,Nox被认为是血管系统中生成ROS的主要氧化酶。
Nox蛋白家族包括Nox 1-5、双功能氧化酶1及双功能氧化酶2几种亚型,其结构由2个组织亚单位p47phox和Noxo1亚基、2种激活亚单位p67phoxa和Noxa1、2个调节亚单位p22phox和p40phox以及结合在Rac蛋白上的调节亚单位组成。Nox蛋白的亚型具有不同的结构,其中Nox1-4蛋白具有60%氨基酸序列的同源性,其N末端有6个保守的跨膜区,也叫跨膜a单环,其C末端包含有NADPH结合位点。Nox5除了具有与Nox1-4相同的基本结构域外,还在其N端有4个被认为是钙离子结合位点的E-螺旋/F-螺旋手结构域。Duox1和Duox2具有与Nox5相似的结构,并且在其N末端有1个髓过氧化物酶样的胞外域。Nox蛋白家族成员中,Nox2最先在中性粒细胞和吞噬细胞内被发现,因此它常被称为吞噬细胞Nox,它的催化亚基gp91Phox和调节亚基p22Phox在细胞膜上形成异二聚体,其他亚基通常位于细胞质,包括p40Phox、p47Phox、p67Phox和小G蛋白Rac。不同的Nox表现出组织分布的特异性并具有不同的功能。脉管系统主要表达Nox1、Nox2、Nox4和Nox5,在血管EC中,ROS主要由Nox2和Nox4产生。
2 ROS与EPC
血管的新生对于创口的愈合起着关键作用,在创口血管的新生过程中,来源于骨髓的EPC的动员有助于受损害创口血管的发生。适当水平的ROS对于造血干细胞的衰老和分化具有重要的影响。更为重要的是,ROS调控的信号路径有助于来源于骨髓的EPC释放。粒细胞集落刺激因子诱导EPC和其他祖细胞的动员过程能够被抗氧化剂N-乙酰-L-半胱氨酸所阻止[4]。有研究表明,在敲除了Nox2基因的后肢缺血老鼠模型中,其缺血诱导的血流恢复减少,EPC的动员受到损害,但是这一损害过程能够通过移植野生型老鼠的EPC而得到改善[5]。在给予低氧或者促红细胞生成素刺激的条件下,敲除了Nox2基因的EPC的动员也是减少的,这可能归因于ROS生成的缺乏,以致于不能激活促红细胞生成素信号路径,从而使得EPC的动员减少。在骨髓中,基质的降解和重塑对于EPC的动员和生长因子的释放十分重要,在骨髓细胞,通过瘦素结合其受体能够激活由Nox2所诱导的ROS的生成[6]。伴随着ROS的生成,基质金属蛋白酶9的表达上调,它能够通过释放水溶性Kit配体从而增强EPC的动员。除此之外,ROS的生成有助于血管细胞黏附分子-1在EC的表达,这样能够促进造血干细胞亚群Lin-Sca-1+cKit+CD34-(LSKCD34-)的募集和增殖[7]。
3 ROS与EC
创口血管的生成过程包括EC的迁移、增殖和基质的重塑,其受氧化还原信号调控。在敲除了Nox2基因的后肢缺血老鼠模型中,其血管生成的能力以及EC的增殖和迁移能力也是减弱的[8]。
3.1 EC的迁移
当受到创口损害的刺激时,EC迁移并且覆盖创口中被损害的血管,这一过程涉及复杂的细胞外基质与细胞、细胞与细胞之间的相互作用。血管内皮钙黏蛋白形成二聚体直接结合于β连环蛋白和p120连环蛋白上,进而与α连环蛋白结合,后者又与肌动蛋白细胞骨架结合形成钙黏蛋白-连环蛋白复合体。在细胞迁移的起初,位于血管壁上处于静止状态的EC首先分解主要由血管内皮钙黏蛋白组成的细胞间连接。IQGAPl(IQ-domain GTPase-activating protein 1)是一种进化保守的多结构域蛋白,它作为一种细胞骨架蛋白,能够结合于肌动蛋白、β连环蛋白、微管正端示踪蛋白CLIP-170、Rac蛋白、Cdc42蛋白、钙调蛋白以及其他与形成细胞骨架相关的蛋白[9]。当EC受到血管内皮生长因子(VEGF)刺激时,IQGAP1蛋白通过激活血管内皮生长因子受体(VEGFR)以及募集Rac1蛋白和Nox亚单位从而产生ROS; IQGAP1蛋白与VEGFR的结合还能够进一步激活c-Src蛋白激酶[10]磷酸化IQGAP1蛋白,募集更多的Nox亚单位或者激活Rac交换因子Vav2,从而增加ROS的生成。
3.2 EC的增殖与生存
增殖的EC能够产生高水平的超氧化物和过氧化氢。通过Nox2和Nox4产生的ROS能够增强EC的增殖和生存能力,这可能与ROS激活酪氨酸激酶受体以及磷酸化p38、ERK和Akt信号通路有关。在血管EC中,Nox2表达的沉默能够诱导凋亡的关键酶半胱天冬酶3和半胱天冬酶7的激活,而在血清剥夺刺激的情况下,Nox4过表达能够抑制凋亡酶的激活,这表明来源于各种Nox亚型的ROS能够发挥抗凋亡效应。在能源不足的条件下,细胞启动通过溶酶体降解细胞质内受损害成分的促生存机制的过程被称之为自噬[11]。它不仅是一种细胞死亡方式,同时也是一种细胞自我保护机制,有利于EC的生存。在EC受到应力刺激时,ROS作为信号分子,已被报道参与细胞内自噬的调节[12]和促进EC的生存[13]。抑制线粒体生成ROS能够减少由趋化素或者2-脱氧-D-葡萄糖诱导EC自噬过程中腺苷酸活化蛋白激酶的激活[14]。此外,在血管生成的过程中,由ROS调节的自噬对于EC的迁移和管状形成很关键[15]。
4 ROS与VSMC
VSMC作为血管壁重要的组成成分,具有收缩和舒张血管、调节血压以及分配血流的功能,有很强的可塑性。当血管受到损害或处于病理状态时,VSMC从静息的可收缩状态向具有迁移和生物合成能力的状态分化[16]。在创口血管的生成过程中,ROS能调节VSMC的分化、增殖、迁移等多种功能。ROS能够通过Nox4、Nrf3、Pla2g7及其他调节蛋白来调控平滑肌细胞(SMC)的分化,还能够增强血管平滑肌分化/收缩状态的维持能力。VSMC在受到血红素[17]和尾加压素Ⅱ[18]刺激时,能激活Nox和ROS的形成,促进SMC的增殖。
在创口血管生成过程中,VSMC需要迁移并且覆盖已存在的侧支血管网,为成熟的血管提供机械支持与收缩力。ROS促进SMC迁移的机制是多样的。首先,ROS激活与迁移相关的信号通路,这些通路包括c-Src-PDK1-PAK[19]与c-Src-EGFR-PI3K/ERK[20]。其次,SMC的迁移需要细胞外基质的降解。ROS能够影响细胞间神经-钙黏蛋白的脱落,这与其影响EC的血管内皮钙黏蛋白相似。细胞外基质的降解通常需要基质金属蛋白酶裂解活性的激活,通过Nox1激活上皮生长因子受体,基质金属蛋白酶9能够裂解神经-钙黏蛋白[21]。创口血管生成时,ROS的生成对于基质金属蛋白酶的转录、激活与合成具有重要的作用[21],从而促进细胞的迁移。基质金属蛋白酶2是一种潜在的促进SMC迁移的亚型。最后,ROS的生成能够进一步调控迁移细胞骨架的重构[22]。在迁移细胞骨架形成过程中,丝切蛋白作为肌动蛋白解聚的调节剂[23],其活性主要通过磷酸化/去磷酸化来调节。丝切蛋白通过LIM激酶磷酸化和失活,通过磷酸酶SSH-1L去磷酸化而复活[24]。Nox1通过氧化14-3-3蛋白生成ROS,进而释放磷酸酶SSH-1L激活丝切蛋白,从而调控迁移细胞的细胞骨架重构[25, 26]。除此之外,VSMC胞内产生的ROS能够通过钙离子的内流增加细胞钙离子的动员,增强其收缩能力[27]。
5 结语
近年来,ROS被越来越广泛地认为是多种正常生理过程或者病理状态下关键的调节剂。创口的愈合过程涉及血管新生,越来越多的研究发现ROS对于创口血管的新生过程是不可或缺的,这一血管新生过程包括EPC的动员和分化,EC和VSMC的迁移、增殖以及分化。在本文中,我们从ROS的来源与Nox家族,ROS与EPC、EC、VSMC等方面讨论了ROS对于创口血管新生过程的影响。尽管如此,当前还有许多关于ROS的信号调节问题以及病理生理功能问题亟待解决,尤其是如何避免细胞内ROS过多蓄积从而造成氧化应激损害。因此,更好地了解ROS在不同病理或生理状态下如何影响血管系统[28],特别是对血管新生过程中所涉及细胞的调控机制有何影响,有助于我们寻找新的治疗途径以调控创口愈合的血管新生过程。
活性氧簇(ROS)是一类氧衍生的具有较强化学反应的活性分子,包括超氧阴离子、过氧化氢、过氧亚硝基阴离子、次氯酸、羟自由基、一氧化氮等。它的生成是多种刺激因素作用的结果,这些刺激因素包括生长因子、细胞因子、低氧诱导因子、剪切应力等。传统观点认为,ROS通过自由基链式反应引起生物体的核酸、脂肪、蛋白质等大分子的损害。近几十年来,大量的研究表明,氧化应激对于血管疾病的发病具有重要的作用,例如与高血压、动脉粥样硬化和再狭窄的发生有着密切的关系。近年的研究也表明,对于多种生理活动过程而言,ROS均具有重要的调节作用,细胞内多种重要信号路径的调节都是由ROS介导的,例如G蛋白偶联受体、Notch路径[1]、Wnt-β-catenin路径[2]、促分裂原活化蛋白激酶路径、JAK-STAT路径、核因子κB(NF-κB)、PI3K/AKT路径等,其调节信号的转导过程被称之为可逆性的氧化还原过程。除此之外,ROS还可以激活转录因子(例如激活蛋白1、NF-κB自身、c-Jun氨基末端激酶等),调控细胞的生长、分化、迁移、凋亡、衰老等多种生理活动。
创口愈合缓慢会增加医疗负担,尤其影响老年、糖尿病及肥胖患者。组织损伤及缺血所引起的血管生长对于创口的愈合非常关键。目前发现存在2种不同形式的血管新生:血管的发生,由骨髓来源分化而成的循环内皮祖细胞(EPC)所形成;血管的生成,由已存在血管局部内皮细胞(EC)的芽生而形成。创口血管的新生包括血管的发生、生成以及血管平滑肌细胞(VSMC)迁移从而形成血管壁的过程[3]。这一过程涉及EPC的动员和分化、EC的迁移、VSMC表型的转化等,ROS在其中的作用不可或缺。因此,现就ROS如何影响创口愈合过程中血管的新生进行综述。
1 ROS的来源与还原型辅酶Ⅱ氧化酶家族(Nox)
ROS来源于包括Nox、黄原呤氧化酶、线粒体呼吸链、非耦合的一氧化氮合成酶、髓过氧化物酶、脂氧合酶、环氧合酶、细胞色素P450和血红素加氧酶的多酶系统。在上述多种能够产生ROS的氧化酶中,Nox被认为是血管系统中生成ROS的主要氧化酶。
Nox蛋白家族包括Nox 1-5、双功能氧化酶1及双功能氧化酶2几种亚型,其结构由2个组织亚单位p47phox和Noxo1亚基、2种激活亚单位p67phoxa和Noxa1、2个调节亚单位p22phox和p40phox以及结合在Rac蛋白上的调节亚单位组成。Nox蛋白的亚型具有不同的结构,其中Nox1-4蛋白具有60%氨基酸序列的同源性,其N末端有6个保守的跨膜区,也叫跨膜a单环,其C末端包含有NADPH结合位点。Nox5除了具有与Nox1-4相同的基本结构域外,还在其N端有4个被认为是钙离子结合位点的E-螺旋/F-螺旋手结构域。Duox1和Duox2具有与Nox5相似的结构,并且在其N末端有1个髓过氧化物酶样的胞外域。Nox蛋白家族成员中,Nox2最先在中性粒细胞和吞噬细胞内被发现,因此它常被称为吞噬细胞Nox,它的催化亚基gp91Phox和调节亚基p22Phox在细胞膜上形成异二聚体,其他亚基通常位于细胞质,包括p40Phox、p47Phox、p67Phox和小G蛋白Rac。不同的Nox表现出组织分布的特异性并具有不同的功能。脉管系统主要表达Nox1、Nox2、Nox4和Nox5,在血管EC中,ROS主要由Nox2和Nox4产生。
2 ROS与EPC
血管的新生对于创口的愈合起着关键作用,在创口血管的新生过程中,来源于骨髓的EPC的动员有助于受损害创口血管的发生。适当水平的ROS对于造血干细胞的衰老和分化具有重要的影响。更为重要的是,ROS调控的信号路径有助于来源于骨髓的EPC释放。粒细胞集落刺激因子诱导EPC和其他祖细胞的动员过程能够被抗氧化剂N-乙酰-L-半胱氨酸所阻止[4]。有研究表明,在敲除了Nox2基因的后肢缺血老鼠模型中,其缺血诱导的血流恢复减少,EPC的动员受到损害,但是这一损害过程能够通过移植野生型老鼠的EPC而得到改善[5]。在给予低氧或者促红细胞生成素刺激的条件下,敲除了Nox2基因的EPC的动员也是减少的,这可能归因于ROS生成的缺乏,以致于不能激活促红细胞生成素信号路径,从而使得EPC的动员减少。在骨髓中,基质的降解和重塑对于EPC的动员和生长因子的释放十分重要,在骨髓细胞,通过瘦素结合其受体能够激活由Nox2所诱导的ROS的生成[6]。伴随着ROS的生成,基质金属蛋白酶9的表达上调,它能够通过释放水溶性Kit配体从而增强EPC的动员。除此之外,ROS的生成有助于血管细胞黏附分子-1在EC的表达,这样能够促进造血干细胞亚群Lin-Sca-1+cKit+CD34-(LSKCD34-)的募集和增殖[7]。
3 ROS与EC
创口血管的生成过程包括EC的迁移、增殖和基质的重塑,其受氧化还原信号调控。在敲除了Nox2基因的后肢缺血老鼠模型中,其血管生成的能力以及EC的增殖和迁移能力也是减弱的[8]。
3.1 EC的迁移
当受到创口损害的刺激时,EC迁移并且覆盖创口中被损害的血管,这一过程涉及复杂的细胞外基质与细胞、细胞与细胞之间的相互作用。血管内皮钙黏蛋白形成二聚体直接结合于β连环蛋白和p120连环蛋白上,进而与α连环蛋白结合,后者又与肌动蛋白细胞骨架结合形成钙黏蛋白-连环蛋白复合体。在细胞迁移的起初,位于血管壁上处于静止状态的EC首先分解主要由血管内皮钙黏蛋白组成的细胞间连接。IQGAPl(IQ-domain GTPase-activating protein 1)是一种进化保守的多结构域蛋白,它作为一种细胞骨架蛋白,能够结合于肌动蛋白、β连环蛋白、微管正端示踪蛋白CLIP-170、Rac蛋白、Cdc42蛋白、钙调蛋白以及其他与形成细胞骨架相关的蛋白[9]。当EC受到血管内皮生长因子(VEGF)刺激时,IQGAP1蛋白通过激活血管内皮生长因子受体(VEGFR)以及募集Rac1蛋白和Nox亚单位从而产生ROS; IQGAP1蛋白与VEGFR的结合还能够进一步激活c-Src蛋白激酶[10]磷酸化IQGAP1蛋白,募集更多的Nox亚单位或者激活Rac交换因子Vav2,从而增加ROS的生成。
3.2 EC的增殖与生存
增殖的EC能够产生高水平的超氧化物和过氧化氢。通过Nox2和Nox4产生的ROS能够增强EC的增殖和生存能力,这可能与ROS激活酪氨酸激酶受体以及磷酸化p38、ERK和Akt信号通路有关。在血管EC中,Nox2表达的沉默能够诱导凋亡的关键酶半胱天冬酶3和半胱天冬酶7的激活,而在血清剥夺刺激的情况下,Nox4过表达能够抑制凋亡酶的激活,这表明来源于各种Nox亚型的ROS能够发挥抗凋亡效应。在能源不足的条件下,细胞启动通过溶酶体降解细胞质内受损害成分的促生存机制的过程被称之为自噬[11]。它不仅是一种细胞死亡方式,同时也是一种细胞自我保护机制,有利于EC的生存。在EC受到应力刺激时,ROS作为信号分子,已被报道参与细胞内自噬的调节[12]和促进EC的生存[13]。抑制线粒体生成ROS能够减少由趋化素或者2-脱氧-D-葡萄糖诱导EC自噬过程中腺苷酸活化蛋白激酶的激活[14]。此外,在血管生成的过程中,由ROS调节的自噬对于EC的迁移和管状形成很关键[15]。
4 ROS与VSMC
VSMC作为血管壁重要的组成成分,具有收缩和舒张血管、调节血压以及分配血流的功能,有很强的可塑性。当血管受到损害或处于病理状态时,VSMC从静息的可收缩状态向具有迁移和生物合成能力的状态分化[16]。在创口血管的生成过程中,ROS能调节VSMC的分化、增殖、迁移等多种功能。ROS能够通过Nox4、Nrf3、Pla2g7及其他调节蛋白来调控平滑肌细胞(SMC)的分化,还能够增强血管平滑肌分化/收缩状态的维持能力。VSMC在受到血红素[17]和尾加压素Ⅱ[18]刺激时,能激活Nox和ROS的形成,促进SMC的增殖。
在创口血管生成过程中,VSMC需要迁移并且覆盖已存在的侧支血管网,为成熟的血管提供机械支持与收缩力。ROS促进SMC迁移的机制是多样的。首先,ROS激活与迁移相关的信号通路,这些通路包括c-Src-PDK1-PAK[19]与c-Src-EGFR-PI3K/ERK[20]。其次,SMC的迁移需要细胞外基质的降解。ROS能够影响细胞间神经-钙黏蛋白的脱落,这与其影响EC的血管内皮钙黏蛋白相似。细胞外基质的降解通常需要基质金属蛋白酶裂解活性的激活,通过Nox1激活上皮生长因子受体,基质金属蛋白酶9能够裂解神经-钙黏蛋白[21]。创口血管生成时,ROS的生成对于基质金属蛋白酶的转录、激活与合成具有重要的作用[21],从而促进细胞的迁移。基质金属蛋白酶2是一种潜在的促进SMC迁移的亚型。最后,ROS的生成能够进一步调控迁移细胞骨架的重构[22]。在迁移细胞骨架形成过程中,丝切蛋白作为肌动蛋白解聚的调节剂[23],其活性主要通过磷酸化/去磷酸化来调节。丝切蛋白通过LIM激酶磷酸化和失活,通过磷酸酶SSH-1L去磷酸化而复活[24]。Nox1通过氧化14-3-3蛋白生成ROS,进而释放磷酸酶SSH-1L激活丝切蛋白,从而调控迁移细胞的细胞骨架重构[25, 26]。除此之外,VSMC胞内产生的ROS能够通过钙离子的内流增加细胞钙离子的动员,增强其收缩能力[27]。
5 结语
近年来,ROS被越来越广泛地认为是多种正常生理过程或者病理状态下关键的调节剂。创口的愈合过程涉及血管新生,越来越多的研究发现ROS对于创口血管的新生过程是不可或缺的,这一血管新生过程包括EPC的动员和分化,EC和VSMC的迁移、增殖以及分化。在本文中,我们从ROS的来源与Nox家族,ROS与EPC、EC、VSMC等方面讨论了ROS对于创口血管新生过程的影响。尽管如此,当前还有许多关于ROS的信号调节问题以及病理生理功能问题亟待解决,尤其是如何避免细胞内ROS过多蓄积从而造成氧化应激损害。因此,更好地了解ROS在不同病理或生理状态下如何影响血管系统[28],特别是对血管新生过程中所涉及细胞的调控机制有何影响,有助于我们寻找新的治疗途径以调控创口愈合的血管新生过程。